 Return to the Histology Tutorial menu.
Return to the Histology Tutorial menu. Return to the Histology Tutorial menu.
Return to the Histology Tutorial menu.|
The female genital tract consists of the external genitalia and the internal genitalia with vagina, uterus, fallopian tubes, and ovaries. External Female GenitaliaThe external genitalia, including labia majora, labia minora, and clitoris, are covered by keratinized stratified squamous epithelium. The clitoris has erectile tissue consisting of irregular endothelial-lined sinuses with intervening fibrovascular connective tissue. The clitoris has numerous sensitive nerve endings. The clitoris and labia minora do not contain fat or muscle or hair follicles. The outer labia have hair follicles. VaginaThe vagina is a muscular tube lined by a stratified squamous mucosa containing abundant glycogen. There is no epithelial keratin layer, but the mucosa is protected by an acid environment (lactic acid) resulting from bacterial growth on a glycogen substrate supplied by the mucosa. There is a connective tissue layer beneath, the lamina propria, which may contain some lymphocytes, with prominent vessels, but no glands are present here. However, there is a 1 cm Bartholin's gland in the vaginal wall that supplies mucinous secretions. Most of the lubrication of the vagina that occurs with sexual arousal comes from transudation of fluid from the lamina propria, mixed with cervical gland secretions. Surrounding smooth muscle bundles are present, mostly longitudinal, with some oblique and circular bundles admixed. The outer adventitia contains nerve plexuses. The lower end of the vagina has a surrounding skeletal muscle--the pubococcygeus. The upper vagina merges with the uterus at the cervix. CervixThe cervix is the lower end of the uterus. In the adult, the cervical and endocervical region comprises 1/3 of the length of the uterus. The outer cervix is lined by a stratified squamous mucosa containing abundant glycogen. The underlying fibrovascular connective tissue of the lamina propria merges with smooth muscle bundles; there may be some scattered lymphocytes. At the cervical os, the squamous epithelium changes to a tall columnar mucinous epithelium. This squamocolumnar junction is called the transformation zone. Beneath the columnar mucosa in the lamina propria are prominent branching tubular glands also lined by columnar mucosa producing mucin. These are endocervical glands that extend into the lower uterine segment along the endocervical canal. EndometriumThe body of the uterus consists of a thick muscular wall surrounding the endometrial cavity, which in the non-pregnant uterus is just a slit-like cavity. The endometrial cavity is lined by the endometrium, which is highly responsive to hormonal changes. The actual endometrial epithelial surface of columnar cells, some of which are ciliated, is not prominent. What is prominent are glands in a thick lamina propria. Thus, endometrium consists of glands and stroma. The stroma is composed of many small fibroblastic cells along with scattered lymphocytes, macrophages, and blood vessels. The endometrium is divided into two indistinct layers: a stratum basalis and a stratum functionalis. The basalis consists of a narrow layer of tubular endomtrial glands surrounded by a dense stroma. The basalis can regenerate the overlying functionalis, which is hormonally responsive and changes with the menstrual cycle. In this cycle, the proliferative phase builds up the endometrium, and the glands appear tubular. In the secretory phase following ovulation, the glands become more tortuous and are filled with secretions. The purpose of these changes is to provide an environment to sustain a fertilized ovum that may implant. If implantation does not occur, the functionalis is shed and the process begins again. If a fertilized ovum implants, then hormonal changes occur that prevent endometrial shedding, and the stroma becomes the "decidua" which initially supports the embryo and then becomes the bed of the placenta. 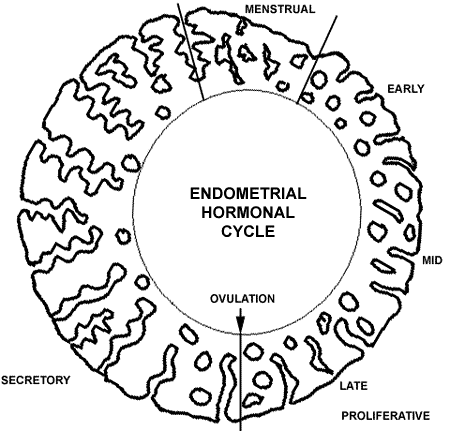
The endometrial hormonal cycle is diagrammed above. The "average" cycle is 28 days. The time from ovulation to menstruation in the secretory portion of the cycle is a constant 14 day period. The menstrual portion of the cycle averages 3 to 7 days. The proliferative portion of the cycle is variable among women, but tends to remain the same for any one person. MyometriumThe myometrium is the thick muscle layer that forms the uterine wall, typically about 1 to 2 cm thick in the non-pregnant uterus. The myometrium is composed of interlacing bundles of smooth muscle. Most of the uterus extends into the lower pelvic part of the peritoneal cavity and is covered by a serosa of mesothelium. Fallopian TubeAlso known as the oviduct, the fallopian tube is a bilateral structure extending from the cornu of the top lateral uterus to the ovary. The fallopian tube has a narrow isthmus near the uterus that widens to an ampullary region just before terminating as abundant loose folds, or fimbria, that lie in close proximity to the ovary. Its purpose is to conduct sperm upward to meet an ovum and then conduct a fertilized ovum distally to implant on the endometrium. The fallopian tube is lined by a columnar epithelium. Some epithelial cells have cilia. Some of the cilia beat upward to help sperm ascend, while other beat downward to conduct ova toward the uterus. Some epithelial cells have a secretory function and are called "peg cells." The secretion products of peg cells function in "capacitation" of sperm which causes them to mature and become capable of fertilizing an ovum lurking in the tube. Beneath this epithelium is a lamina propria and then a muscularis of inner circular and outer longitudinal layers. The structure of the fallopian tube is complex in cross-section, with many fingers of epithelial-covered lamina propria creating a maze-like appearance. There is a surrounding mesothelium. OvaryEach ovary contains the ova, or primordial germ cells, that will eventually contain a haploid number of chromosomes. Thus, an ovum has a karyotype of 23, X and can be fertilized by either a 23, X or 23, Y sperm to produce a 46, XX female or 46, XY male offspring. The fetal ovary contains mostly ova, with an indistinct intervening stroma. Beginning in late gestation and throughout childhood, ova disappear, until at the time of reproductive maturity at menarche, less than a couple of hundred ova remain in each ovary which can be released during each menstrual cycle throughout the next 30 or so reproductive years. The adult ovary consists of a cortex and a medulla. A mesothelium, also known as the germinal epithelium, surrounds the ovary. the outer cortex consists mainly of a stroma, or interstitium, composed of small fusiform cells that can transform under hormonal influence to support the developing ova. the cortex contains scattered ova. A primordial follicle consists of just the oocyte surrounded by a flattened layer of stromal cells. The central medullary portion of the ovary contains abundant blood vessels in connective tissue. Both the ovary and the fallopian tube are supported and held in place by a broad ligament of connective tissue with the vascular supply. There are a few scattered hilus cells (the female equivalent of testicular Leydig cells) capable of secreting androgenic steroids. During reproductive years, primordial follicles are transforming into primary follicles that have a layer of granulosa cells to support the oocyte. A primary follicle is surrounded by interstitial cells termed the theca. As this structure enlarges and fluid accumulates centrally, it becomes a secondary follicle. A mature follicle takes about 10 to 14 days to develop, at which time ovulation with release of the oocyte can take place. At ovulation, the fimbriated end of the fallopian tube surrounds the ovary and ciliary action draws the released ovum into the fallopian tube. After ovulation, the mature follicle that released the oocyte becomes a corpus luteum. If the oocyte becomes fertilized by the sperm and implants in the endometrium, hormonal changes transform the corpus luteum into a steroid factory, producing much progesterone, that maintains the pregnancy. If fertilization does not occur, the corpus luteum involutes into a corpus albicans that appears as a cloud-like pink scar. At menopause the ovary loses regular hormonal stimulation and atrophies. In the postmenopausal ovary, there are no follicles, only a dense stroma containing corpora albicantia, and a medulla with thick, hyalinized blood vessels. Female BreastEach of the two breasts is essentially a set of modified sweat glands designed to produce milk for a baby following delivery from a pregnancy. The female breast is composed of glands with ducts and lobules in a fibrofatty stroma. The female breasts increase in size at puberty. Breast size is determined more by the amount of adipose tissue than by the amount of glands. Occasionally, breast development is not symmetrical at puberty, and cosmetic surgery can be performed to remove some breast tissue on one side, but functionality is maintained. Breast augmentation with implants occurs beneath the breast tissue on the chest wall to preserve functionality, though radiologic mammographic studies are compromised. The breast is composed of lobules that are normally small and are not producing secretions. They drain to ducts lined by a stratified squamous epithelium. The duct systems eventually coalesce to form large lactiferous sinuses that drain to the nipple. Each lobule is surrounded by myoepithelial cells that can contract under the influence of oxytocin. The breasts will enlarge somewhat in the latter part of each menstrual cycle, but when pregancy occurs, the lobules become larger and more numerous. After birth, secretions consist of pinched off portions of lobular epithelium (making the milk rich in lipid). Suckling by the baby on the nipple produces release of oxytocin to contract myoepithelial cells so that milk is expressed. As long as there is suckling from the baby, the breast will continue to produce milk, and a hormonal millieu is maintained that prevents a return to regular menstrual cycles. This is the principle behind use of a "wet nurse" to provide milk to babies whose natural mothers had died or who could not produce sufficient milk. This ancient practice is important in places where you cannot buy milk from a store to feed babies, or if the baby cannot tolerate a formula that consists of milk from another mammalian species (usually bovine). PlacentaThe placenta is actually a fetal structure derived from the developing embryo. The placenta is a structure implanting into the decidualized secretory endometrium. The placenta actually invades the endometrium to establish a maternal-fetal connection. The cells known as the intermediate trophoblast perform this function. The basic unit of the placenta is a chorionic villus. In the first trimester of pregnancy, the chorionic villi have a central core of loose connective tissue and indistinct vessels surrounded by two layers: the cytotrophoblast and outer syncytiotrophoblast. The villi produce human chorionic gonadotropin (HCG) which helps to maintain the pregnancy. In the second trimester, the syncytiotrophoblast draws up into small "knots" of cells. Intervillous fibrin deposits appear scattered between the villi. The size of the villi decreases. In the late second and in the third trimester, the villi further decrease in size and the vessels within them becomes very prominent. External to the villi are sinuses that are filled with maternal blood. Thus, nutrient and gas exchange occurs across the villi. The vessels from the chorionic villi coalesce into larger vessels that eventually course over the fetal surface of the placenta and join the umbilical cord that connects the placenta to the fetus. The developing fetus is suspended in amniotic fluid (derived from fetal urine) that is contained in the amniotic sac, a structure bounded by the fetal membranes that extend from the placenta to surround the fetus. |

 |